
第三纪
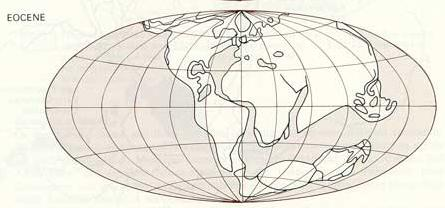
第三纪(始新世)地球
第三纪TertiaryPeriod, 新生代的最老的一个纪(距今6500万年~距今180万年).第三纪的重要生物类别是被子植物、哺乳动物、鸟类、真骨鱼类、双壳类、腹足类、有孔虫等,这与中生代的生物界面貌迥异,标志着“现代生物时代”的来临。
第三纪-梗概
第三纪名称的意思是“第三个衍生期”,这是延用十九世纪对地层划分为四个大时期的分类命名。
1833年,英国C.莱伊尔在研究法国 巴黎盆地软体动物化石时,发现地层越新,软体动物与现代种属相同的越多。他根据地层中含有现代种属的百分比,将第三纪划分为始新世、中新世和上新世。1854年,E.贝利希在德国发现早于中新世的沉积物,提出渐新世。1853年,M.赫奈斯依生物群和沉积物的相似性,将中新世和上新世合称晚第三纪;1866年,K.F.瑙曼则把渐新世和始新世合称早第三纪。1874年,W.P.夏姆珀根据巴黎盆地发现的植物化石,他认为这些化石的层位早于始新世,而称之为古新世。因此,第三纪划分为早第三纪和晚第三纪。早第三纪包括古新世、始新世和渐新世,晚第三纪包括中新世和上新世。早、晚第三纪的分界线约在2330万年前。
习惯上将这个纪分为5个持续时间不等的世,从老到新为:古新世、始新世、渐新世、中新世和上新世。它们的名称是根据现代海洋无脊椎生物种属在第三纪化石中占的比例命名的。因此,根据提出这个系统的查尔斯•莱伊尔爵士的意见,始新世(意思是“现代的拂晓”)有比更新世(译为更为现代)较少的现代物种。在另一种分类中,将第三纪看作一个代,其中的前三个世构成老第三纪而后两个构成新第三纪。
第三纪时被子植物极度繁盛。除松柏类尚占重要地位外,其余的裸子植物均趋衰退。蕨类植物也大大减少且分布多限于温暖地区。第三纪的植物有明显的分区现象,地层中还有许多微体水生藻类化石。
脊椎动物的变化主要表现在爬行动物的衰亡,哺乳类、鸟类和真骨鱼类取而代之,兴起且高度繁盛。第三纪的早期,仍生活着古老、原始的哺乳动物;到了中期,现代哺乳动物的祖先先后出现,逐渐代替了古老、原始的哺乳动物;第三纪晚期,现代哺乳动物群逐渐形成,更是偶蹄类和长鼻类繁盛的时期。尤其马的进化很快。
中生代末,海生无脊椎动物有明显的兴衰现象。盛极一时的菊石类完全绝灭,箭石类极度衰退,而双壳类、腹足类、有孔虫、六射珊瑚、海胆、苔藓虫等则进一步繁盛。第三纪出现的有孔虫分布广泛、进化迅速,对于海相第三系的划分与对比很有意义。此外,它们的生态分带已应用于确定沉积盆地水深的变化。原生动物中的放射虫在第三纪也十分繁盛,在深海研究中占有突出地位。双壳类在第三纪有很大发展,腹足类在第三纪进入极盛期。
陆生的无脊椎动物以双壳类、腹足类、介形类为主,可以根据它们不同时期组合面貌的变化,进行陆相第三系的划分。

第三纪化石
第三纪-地层构造
早第三纪(开始于6500万年前,结束于2330万年前)
早第三纪在地史上经历的时间相对较短,但在这段时间形成的沉积物与中生代有明显的不同。下第三系含有丰富的各种生物类群的。这些化石为下第三系的划分、对比以及标准层序和层型剖面的建立起着关键性的作用。世界上不少地区在早第三纪形成了界线较为分明的海相和海陆过渡相的地层,如西欧下第三系标准层序划分为11个阶。在北美中西部、亚洲中部等地区也形成了以河湖相为主的陆相沉积,尤以美国落基山脉的盆地分布区最为典型。
下第三系除以岩石地层、古地磁和同位素测定为依据外,主要采用古生物组合(或动物群)、层序和层型剖面等对比方法来划分。由于各门类生物进化速率不同,各地沉积并非同时,因此地层的划分常与上述研究提供的论证不相吻合,以致对比和界线常引起争议。如海相地层中的达宁阶(丹麦)和阿基坦阶(法国)的时代归属就有不同认识。有人认为达宁阶的棘皮动物(Echioids)等具有晚白垩纪的特征,但是据克里米亚、高加索和里海周围地区以及丹麦、瑞典南部浮游有孔虫和棘皮动物化石的研究,它们与古新世类型有密切关系。因此较多的学者认为,中生界与新生界分界应置于马斯特里赫阶与达宁阶之间。但仍有一些学者认为,中生界与新生界界线应划在达宁阶与蒙蒂阶之间。阿基坦阶的归属问题,古生物学家从软体动物和哺乳动物研究认为,它的生物群与下部斯坦普阶的化石有着明显的继承关系,而与晚第三纪相比有一定的差异,他们主张阿基坦阶定为渐新世的顶界。但是阿基坦阶的海相微体化石与早中新世的波尔多阶的较近,它又代表了欧洲一次大的海侵时期,因此它作为新第三纪的底界是适当的。不少学者主张下第三系与上第三系分界应置于恰特阶与阿基坦阶间。相比之下,陆相下第三系的划分标准较为一致,一般与美国经典地区的层序为准。下第三系底界为贝尔卡阶之底,它标志了恐龙的消失,哺乳动物有胎盘类开始兴起;上界为维特奈阶之顶,它标志了哺乳动物群开始现代化的特点。
中国下第三系发育齐全。在全国数百个盆地中,广布了很有特色的陆相地层。目前世界上古新统的发现为数不多,主要以美国居多。亚洲在20世纪初仅在蒙古发现了一个地点。1960年以来,中国在8个省和自治区陆续发现了不少重要的古新统地点。中国陆相古新统以下部的上湖组和上部的池江组为代表。古新统已发现的动物化石几百种,其中含有脊椎动物100多种。始新统在中国有良好的发育,如内蒙古地区的典型层序阿尔丁曼阶和锡拉木仑阶已获国际上公认。渐新统的系统研究起步较晚,近年来在内蒙古、宁夏、新疆、广西、云南等地已有较多的发现,这为健全渐新统的层序提供了良好的基础。中国陆相下第三系的下界大致相当含阶齿兽(Bem-lmbd)动物群的上湖组和望虎墩组之底;上界可能相当含副巨犀(Prcertherium)-塔塔鼠(Ttromys)动物群的层位,如内蒙古伊克布拉格组之顶。

第三纪遗留的地貌
中国海相下第三系见于喜马拉雅山区、塔里木西缘和东海陆架──台湾岛。喜马拉雅山区有特提斯型古新统和始新统,古新统包括宗浦组下部,下、中始新统包括宗浦组上部和遮普惹组,它们含大有孔虫、介形虫和软体动物化石。塔里木西缘下第三系主要是喀什群,包括古新统、始新统,另外,渐新统包含于乌恰群底部,它们含有孔虫和软体动物等化石,属于中亚副特提斯海区。台湾北港地区钻孔中发现古新统云林组,有钙质超微化石,西部山麓带和中央山脉带有始新世毕禄山组、十八重溪组、达见组,晚始新世―早渐新世西村组,早渐新世四棱组,中、晚渐新世水长流组、花港组和五指山组,它们主要以有孔虫、钙质超微化石和软体动物化石为时代依据。东海陆架下第三系发育良好,其中古新世灵峰组、始新世瓯江组有孔虫、钙质超微化石序列完整,为西太平洋地区最完善的海相地层序列。

早第三纪地层考察(南宁)
此外,为了使典型层序逐步完善和具有一定的代表性,仍需加强界线和全球性事件的研究。早第三纪地层对比简表(见表[早第三纪地层对比简表]),其中有些地层单元的对比关系尚不够了解,如海相与陆相有关层位的对比等,这在阐明区域或国际间有关层位相互关系带来一定的困难。

早第三纪地层对比

晚第三纪(距今2330万年~距今164万年)
晚第三纪地层的划分与对比有其独特而复杂的历史。

第三纪地层
统的划分
上第三系分为中新统和上新统。这两个统分别源于莱伊尔的地质年代单元,即中新世和上新世。它们是根据地中海周围海相双壳类化石组合中现生种的含量确定的:中新世含现生种18%;上新世含现生种49%。莱伊尔定名时,没有指定典型层位和地点,也没有确定界线,更没有考虑到它们是否能在全球适用。这是造成后来长期争论和混乱的根本原因。虽然如此,绝大多数地质学家都同意,上第三系的统的划分应以研究历史最久的地中海周围的海相地层为准。
中新统的底界,即上第三系的底界应以阿基坦阶的底界为准,它代表了渐新世末海退之后的一次广泛海侵的开始,大约发生于2400万年前。中新统的上限,即上新统的下限应在墨西拿阶和赞克尔阶之间。墨西拿阶代表古地中海区域一次广泛的海退,是有名的成盐期。赞克尔阶代表一次海侵。这一条界线,由于长期来被错误地与陆相地层中的三趾马动物群的出现联系在一起,而推定为距今约1200万年(这也是古哺乳动物学家根据生物演化规律而推算出的结果)。现在已有大量的证据证明对海相中、上新统的界线年龄的推断是错误的。实际它距今只有大约550万年。上新统的顶界,即上第三系的顶界在皮亚琴察阶和卡拉布里亚阶之间。第18届国际地质大会(1948)把这条界线定为上第三系与第四系在海相层中的界线。目前关于这一界线的年代的意见已趋于一致,即生物群总体面貌发生较大变化的界面与古地磁年表中松山反向期中的奥杜威正向亚期的上界大体同时,即距今约170万年。中、上新统在时限上太不均衡,上新统的时限太短(约300万年),而中新统的时限又太长(约1900万年)。此外,这些统的界线不能反映全球性重大的地质和生物事件。事实上,各大陆的晚第三纪的生物和地层年代划分的界线大多与上述界线不相吻合。因此,这种统(世)的划分已失去其作为全球性划分单元的性质。但是,至今也未能提出一个可被普遍接受的方案,当前只能遵循尊重传统的原则,把建立在地中海区域海相地层基础上的中新统和上新统作为国际对比标准。
阶的划分
上第三系的阶只有地区意义。在古地中海周围的海相地层中,中新统分为6个阶,上新统只有两个阶。在中欧和东欧,上第三系主要以副地中海的海相、半咸水封闭内陆海和陆相互层为代表。这些地层在19世纪中期和后期曾是着重研究的对象,出现过大量的地层名称。其中有些是相当著名并被广泛应用的,例如蓬蒂阶等。副地中海有极为复杂的演变历史。因此,它的地层单位之间以及它们和古地中海海相阶的对比造成过极大的混乱。从20世纪60年代末开始,经过多学科长年大量的工作,这些问题已逐步得到澄清。在地中海晚第三纪地层分委员会第7次(1979,雅典)和第8次(1985,布达佩斯)大会上已经提出了中副地中海和东副地中海两个区域地层划分方案,地层对比问题也已基本解决。
在欧洲,陆相上第三系也很发育,而且富含哺乳动物化石。但是陆相地层相变大,连续沉积少,层序关系常常不清(例如裂隙堆积),与海相交互沉积少,多富集于少数层位和地点,这就使海、陆相的对比十分困难。在相当长的一段时期内,欧洲的陆相上第三系没单独的阶。古生物学家越来越感到有建立独立的陆相地层划分顺序的必要。西班牙有较连续的陆相沉积,这一工作首先由西班牙人M.克鲁撒冯开始,于20世纪50年代末建立了瓦里西和土洛里两个阶。1975年,西欧一些古哺乳动物学家在德国慕尼黑聚会,提出了以7个“期”为基础的欧洲陆相地层划分方案。把它们主要称作“期”,而不是“阶”,是因为这些单元大多都不是建立在地层实体的划分上,而是建立在不同的哺乳动物组合上。在这次会上以P.曼为首的一些古生物学家,仿效近年研究洋底钻芯微体古生物学家的办法,把晚第三纪的哺乳动物分成17个带,称作MN带。每一个带都列出了代表性动物群名称,特征属种,首次和末次出现的分子等。这两种划分已经成为当前陆相地层的全球性对比的标准。
北美的上第三系以陆相为主,从一开始就是以哺乳动物的演化为基础划分的。1941年,伍德委员会正式确立了一个划分为6个哺乳动物期的北美上第三系的划分方案。这一方案一直沿用至今,只是这些期的内涵、界限及与欧洲的对比都已经大大前进了。非洲,除地中海沿岸海相地层沿用经典的阶和统的名称外,内陆陆相地层还没有正式建阶,还处在资料积累阶段。印巴次大陆的上第三系以喜马拉雅山南麓巨厚磨拉石相的西瓦利克岩系为代表。在20世纪初,在岩性基础上建立的阶已经提出,但是它们和欧洲划分的对比关系,直到近年才确定下来。

第三纪地层
中国晚第三纪地层
上第三系广布全国,绝大部分属陆相沉积。海相沉积仅分布在台湾、海南岛及沿海极个别的地段。中国上第三系研究起步晚,从20世纪20年代才开始,而且主要限于晚第三纪中后期的哺乳动物化石方面。在30~40年代,曾发表大量有关三趾马动物群方面的专著。但在地层划分方面工作不多。80年代以前,中国一直没有一个独立的上第三系阶一级的划分体系;三趾马动物群和泥河湾动物群分别借用了欧洲的蓬蒂期和维拉弗朗期的名称;其他动物群只是笼统地划归中新世或上新世;而且一直没有发现早中新世的哺乳动物。1984年,李传夔等首次提出一个与欧洲大体对应的7个期(阶)的划分方案。最近的工作表明,中国至少也能划分出15个哺乳动物带来(见表[中国晚第三纪哺乳类层序及与西欧对比])。中国下中新统目前仅发现于青海西宁、甘肃 兰州、河西走廊、新疆准噶尔盆地等处,主要为河湖相碎屑沉积;中中新统发现于宁夏同心、陕西蓝田、江苏泗洪、内蒙古通古尔、山东山旺和云南开远等地。大部分属河湖相沉积,也有部分湖沼沉积。晚中新统广布全国,只华南和东北尚未发现哺乳动物化石。黄河以北、汾河以西主要为土状堆积,即所谓的三趾马红土,汾河以东为河湖相沉积;黄河与长江之间发现较少,在郑州和襄樊一带有灰岩;云南则多沼泽相,有褐煤。70年代在西藏希夏邦马峰北坡也发现三趾马化石。上新统发现较少,仅零散分布于陕西、山西、河北等省和盆地中。
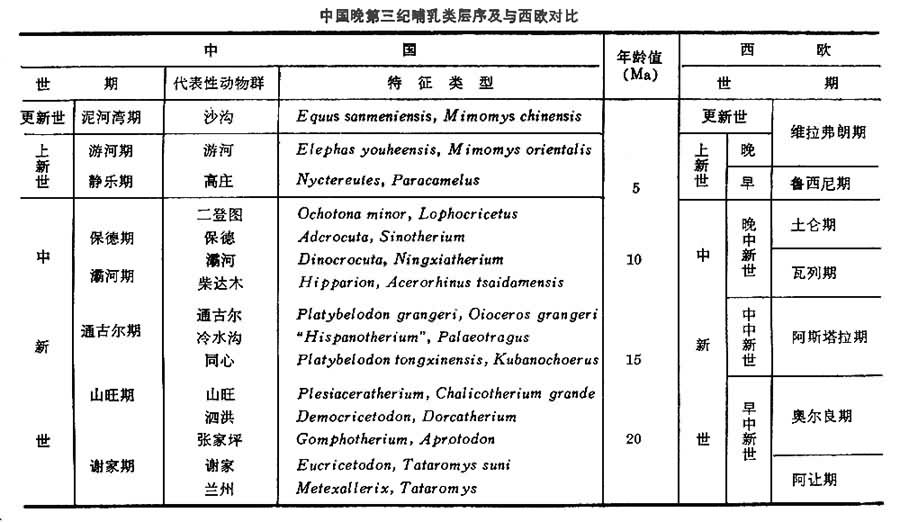
中国晚第三纪哺乳类层序及与西欧对比
第三纪-地史特征
早第三纪由于海底扩张、古陆解体,世界上整个地质构造格局、古地理环境产生了重大的变化。早第三纪时,古地中海最终消失,亚洲大陆最后形成,青藏高原的升起,阿尔卑斯山、喜马拉雅山、落基山和安第斯山等现代山系相继形成,吐尔盖海峡和巴拿马地峡的出现或消失,气候分带渐趋明显,地球上的自然环境向着多样性发展。

形成于第三纪的阿尔卑斯山
晚第三纪全球海、陆轮廓已很接近于现今。整体说来,海洋所占面积较大,而陆地面积较小。现地中海沿岸,如北非阿特拉斯山区、意大利大部、法国和西班牙的地中海沿岸区都曾为古地中海区,法国的西海岸、北欧地区则被大西洋所占;在北美西海岸南部以及墨西哥湾滨海区也被海洋所占。但是,中国的情况不同,中国的东海岸在晚第三纪的大部分时间里更向东扩展,特别是渤海和黄海,大部分还都是陆地。当然最大的不同是副地中海的存在。副地中海是在阿尔卑斯山脉从水下升起后残留在山脉以北部分的水体。水体最大时东西长约5000公里,面积为现地中海的一倍半,从法国罗讷河谷开始,沿阿尔卑斯北麓向东延伸,至少延伸至咸海一带。除罗讷河谷外,副地中海在不同的时期曾通过南斯拉夫萨格勒布低地与亚得里亚海,通过希腊与爱琴海和通过伊朗西北部与波斯湾及印度洋相连通。在中中新世末,约在1300万年前,基本与其他海洋隔绝,变成内陆海,几经分隔,剩下今日的咸海、里海、黑海和匈牙利的巴拉顿湖。
晚第三纪是重要的山脉形成期。现今地球上较高的山脉都是这一时期形成的,如欧洲的阿尔卑斯山、非洲的阿特拉斯山,亚洲的喜马拉雅山,北美的落基山和南美的安第斯山等;一些老的夷平山地,如天山、秦岭等也有活化抬升现象。地壳运动也引起了普遍的火山喷溢。太平洋东、西两岸有大量玄武岩流。在中国境内的有汉诺坝玄武岩,东北地区大片出露,山东山旺和浙江嵊县的玄武岩等。地壳运动也使欧亚大陆与非洲和北美数次分合,形成多次动物群交流。南、北美洲也在上新世末相连过。
晚第三纪开始时,气候由凉爽向湿热转变,中中新世以后,西藏高原的升起无疑对全球的气候产生重大影响。在中国的表现则是气候分带日益明显,并逐渐转向干旱凉爽。
第三纪-气候条件
早第三纪由于各大陆的位置与现代不同,板块漂移、古纬度变化、气温与近今也不同。白垩纪后期到古新世北半球年平均气温曾下降3℃左右;到古新世后期,气温一度回升;而始新世晚期气温又有下降趋势,直至以后才逐渐回升到原有水平。在早第三纪晚期,中国气候有明显的南北分带,如南北为两个潮湿带,中部为干旱带。气候带的展布与今日的纬度斜交。
晚第三纪开始时,气候由凉爽向湿热转变。中中新世后,西藏高原的抬升对全球气候产生重大影响。中国的气候分带日益明显,并逐渐转向干旱凉爽。
总之.第三纪的气候比起现在的气候来,更为温暖、湿润而且较少变化。在第三纪早期,热带和亚热带气候远远延及加拿大北部边界,在稍后的时期内大平原地区则呈现出干旱境况。趋近第三纪末,气候逐渐变冷,预示着更新世最早的冰期即将来临。
在第三纪,中欧、北非和北美东部墨西哥湾沿岸属于热带气候,而北非、西亚、中亚和中国华南大部地区,则属于干燥气候带;东北亚、西西伯利亚、中欧、北美西北部、中国东北地区以及澳大利亚东南部和南美南部,属于温凉潮湿气候带。

第三纪化石
第三纪-生物界
随着白垩纪结束,气候显著的变化给植物生命带来了转机,在景观上增加了多样性。被子植物在早白垩世时出现,到早第三纪极度繁盛。以前由古代羊齿和各种松柏组成的植被逐渐为被子植物所替代。这时的被子植物基本上是乔木,无论从新科、新属,还是从个体数量较之早白垩世有很大增加,整个植物群面貌有了较大的改观。热带、亚热带植物如棕榈等远远深入到北极地区。显花植物和草类的发展给动物界(如昆虫、脊椎动物)的繁荣,创造了必要的条件。
大陆漂移、气候的分带和气温变化,对生命的进化、发展和生态环境产生深远的影响。动物界有海生、陆生和适应半咸水的种类。其中以脊椎动物的变化最为突出。中生代晚期分布广泛的海生爬行动物,如蛇颈龙类、海生蜥蜴类等,在早第三纪被小型鲸类、海狮等海生哺乳类替代。在陆上,中生代占统治地位的恐龙基本消失;而中生代不占突出地位的哺乳动物得到了迅速繁衍,它们占有爬行动物以往的各种生态领域。各大陆和岛屿上充满了各种接近现代面貌的鸟类。从中生代爬行动物占优势过渡到早第三纪哺乳动物的繁盛,这是地球上生命历史变革中的一件大事。关于恐龙为什么大量毁灭仍是一个谜。有的科学家认为是一颗彗星或其他星体陨落在地球上,导致气候骤变或者引起大量酸雨的降落,造成群体动物──恐龙的灭绝。另一些科学家认为是火山喷发、气候变化导致植物界骤变,引起物种的灭绝。
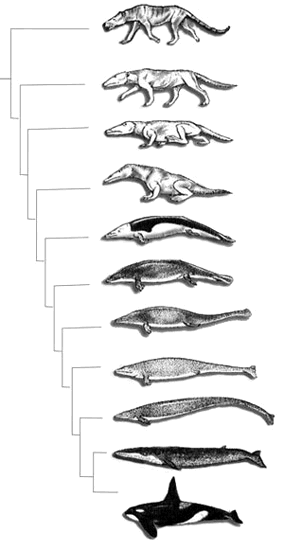
第三纪动物
早第三纪是哺乳动物历史上一个很重要繁衍和进化时期。在晚白垩世时,哺乳动物共有10多个科,但古新世时却增加到40多个科。这些哺乳动物除白垩纪已有的多瘤齿兽目、食虫目和踝节目外,更重要的是各种古老和土著类型的有胎盘类大量发展和分化。它们绝大部分与现代哺乳动物各目都没有直接的系统关系;有许多种类是离奇的或特化的。始新世的哺乳动物科一级的总数比古新世约增加80%;奇蹄目和偶蹄目两大现代目是动物群中的重要角色。此外,啮形动物、食肉目也有了较大的发展和繁衍。渐新世哺乳动物群具有更近代化的面貌。它显示了过渡的特点,一方面是“古老”或“土著”类群的大量衰退、绝灭;另外是近代类群的兴起和开始发展。其他脊椎动物如软骨鱼类、硬骨鱼类、鳄、龟鳖类和蜥蜴类在早第三纪也是常见的化石;鸟类化石也时有发现。至于无脊椎动物,其中双壳类如牡蛎、蚶、海扇和珠蚌异常繁盛;腹足类属种繁多,分布广泛;介形类主要有浪花介、土菱子介、金星介和达尔文介等。叶肢介属种单调,主要是雕饰叶肢介、狭叶肢介等,已处于衰落时期;有孔虫和棘皮动物在世界上分布很广,大有孔虫主要有圆盘虫类、货币虫(Nummulites)类等。由于古地中海区海相早第三系中广布大型原生动物货币虫,故在欧洲常称早第三纪为货币虫纪。而昆虫化石群基本面貌与近代昆虫相似。
早第三纪初期,由于海底扩张,北大西洋断裂切开把北大西洋与北极海相通,当时海生动物大致形成北海、地中海和印度-太平洋3个主要分布中心。而陆上生物因大陆之间像现代这样的连接形式在早第三纪时尚未完成,它的传播与现代仍有明显的不同。根据对脊椎动物的研究,当时的动物地理区大致分为如下区域:古北区,包括欧洲、地中海区域、亚洲中部和北部;新北区,包括北美洲;新热带区,包括南美洲和中美洲;埃塞俄比亚区,包括非洲大部分;澳洲区,包括澳大利亚、新西兰和邻近的一些岛屿;东方区,包括东印度和南亚。(见彩图[第三纪植物群示意图])

第三纪植物群示意图
晚第三纪植物界已基本上由现生属组成,并有大量现生种,只是北半球的喜暖植物的分布更向北延伸。晚第三纪初期北欧还有热带常绿植物,西伯利亚还有温带阔叶林。至上新世末喜暖植物明显南退。
无脊椎动物中大量属种也是现生的。一些早第三纪特有的门类,最著名的如货币虫等,已完全灭绝。双壳类、腹足类和介形类大量衍生。海洋中各种原生动物,如有孔虫、放射虫等极为繁盛,在海相地层划分中起着非常重要的作用。哺乳动物中,欧亚大陆早第三纪的特征门类,如有袋类、肉齿类,奇蹄目中的雷兽、原始貘类、两栖犀、巨犀,偶蹄目中的石炭兽、古猪兽等,除少数残存至中新世初期外,均相继灭绝。代之而起的是长鼻目、肉食目中的鬣狗科、熊科、奇蹄目中的安琪马和三趾马,犀中的板齿犀、大唇犀,偶蹄目中的猪类,反刍动物中的鹿、羊、长颈鹿,以及啮齿目、兔形目中的大量属种。其中绝大部分都是灭绝的种和属。
根据哺乳动物的状况,晚第三纪可划分为4个时期:早中新世是残存的和高度特化的早第三纪分子和少量晚第三纪分子的时期;中中新世,是安琪马动物群时期,此时长鼻目自非洲、安琪马自北美迁入欧亚形成一个全新的动物群;晚中新世至早上新世,为三趾马动物群时期,三趾马从北美迁入,大批草原型动物大量出现;晚上新世,真象从非洲,真马从北美迁入欧亚大陆形成另一个动物群,并逐步演化为今日之动物群。非洲的动物群和欧亚大陆的很接近,只是某些类别的祖先类型(长鼻类、蹄兔类等)出现更早,而有些门类则延续时间更长(如肉齿类等)。北美和欧亚大陆只是间断联接,动物群差别较大,属不同动物区系。至于南美洲和大洋洲,差别就更大了。
植物界的地理分区,中新世已较明显。在中国至少可以分成4个区:华北区、华东沿海区、西藏高原区和西北区。哺乳动物的分区不明显。中国古北区和东洋区的区分约始于晚中新世,而且这条界线可能在长江以南。以淮河和秦岭为界的古北区和东洋区大概形成于上新世。(见彩图[第三纪动物群示意图])

第三纪-矿

铁矿石
早第三纪由于沉积环境多样化,在中国不仅形成了巨厚的磨拉石建造,而且在广大地区形成了各种成因类型的陆相、海相、海陆过渡相的沉积物。这些沉积物中蕴藏的资源相当丰富,主要有含铜砂岩和各种盐类。生成石油的原始物质是有机物。由于早第三纪有大量动植物遗体的堆积,不仅在海相地层而且在陆相地层中都有油田的存在。石油主要见于长期下沉的拗陷和大中型湖泊沉积盆地,如早第三纪初、中期有关层位中。煤受一定的构造和气候因素控制,常见于暖温(亚热带)地区,因气候温暖潮湿适宜于煤的形成和聚集。早第三纪是一个重要的成煤时期,在中国主要见于秦岭以北,贺兰山-六盘山以东地区和南岭以南珠江-右江地区。同时含煤层位中大都含有油页岩层,如中国抚顺、茂名均见有较厚的油页岩层。早第三纪盐类沉积相当丰富,主要有石膏、岩盐、芒硝、天然碱和钾盐等。盐类的形成除具备深坳而封闭较好的盆地外,更重要的是要具有较长期干旱气候的环境。早第三纪时,中国干旱带占有相当大的地区,是盐类富集的天然场所。此外,还有含铜砂岩、铀的氧化物等沉积。
上第三系形成的最主要的矿产是油气。地中海和北美东、西海岸的大油田有相当一部分是晚第三纪形成的。地中海地区麦兴阶是重要的膏盐层。此外,还有褐煤、残积型铁矿、锰矿和硅藻土矿等。
第三纪-参考资料
1.中国百科 http://www.chinabaike.com/article/316/327/2007/2007022052343.html
http://www.chinabaike.com/article/316/327/2007/2007022052235.html
2.百度百科 http://baike.baidu.com/view/66051.htm
3.李云通等:中国的第三系,《中国地层:13》,地质出版社,北京,1984。
